

RF-EMF: Three Biological Pathways, One Upstream Driver
Part 6. Different pathways disturb different oscillators but ultimately affect the same biological timing network.

Please note: This is a complex topic. If any factual errors, omissions, or inaccuracies are identified, please contact PSGRNZ so that corrections or clarifications can be considered.
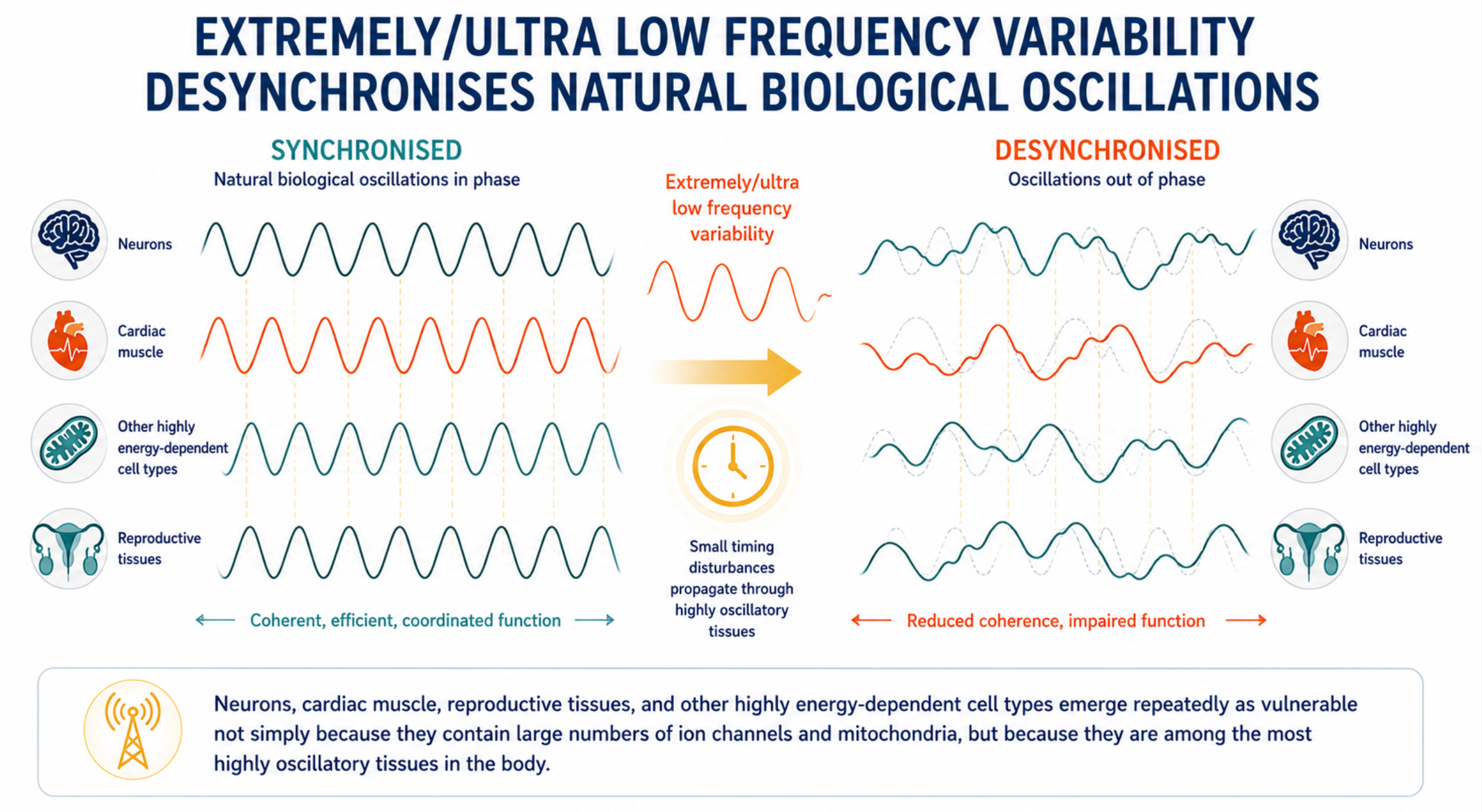
Biological vulnerability is not primarily about energy absorption; it is about disruption of timing fidelity in systems that depend on tightly regulated oscillatory behaviour.
The cells and tissues most consistently damaged by non-thermal electromagnetic field exposure, neurons, cardiac muscle, ovarian granulosa cells, and spermatogenic cells, share two features: exceptionally high voltage-gated ion channel density and exceptionally high mitochondrial load. These exact properties make a cell vulnerable to the two physical transduction mechanisms through which extremely low frequency (ELF)-modulated wireless signals couple into biology.
A useful way to understand this vulnerability is to recognise that living systems are organised around nested biological oscillators operating across many timescales. Electrical membrane potentials fluctuate in milliseconds, calcium signals oscillate over milliseconds to seconds, mitochondrial redox systems generate rhythmic bursts of reactive oxygen species, neuronal networks synchronise across frequency bands, hormones are released in pulses, and circadian clocks coordinate physiology over twenty-four hours.
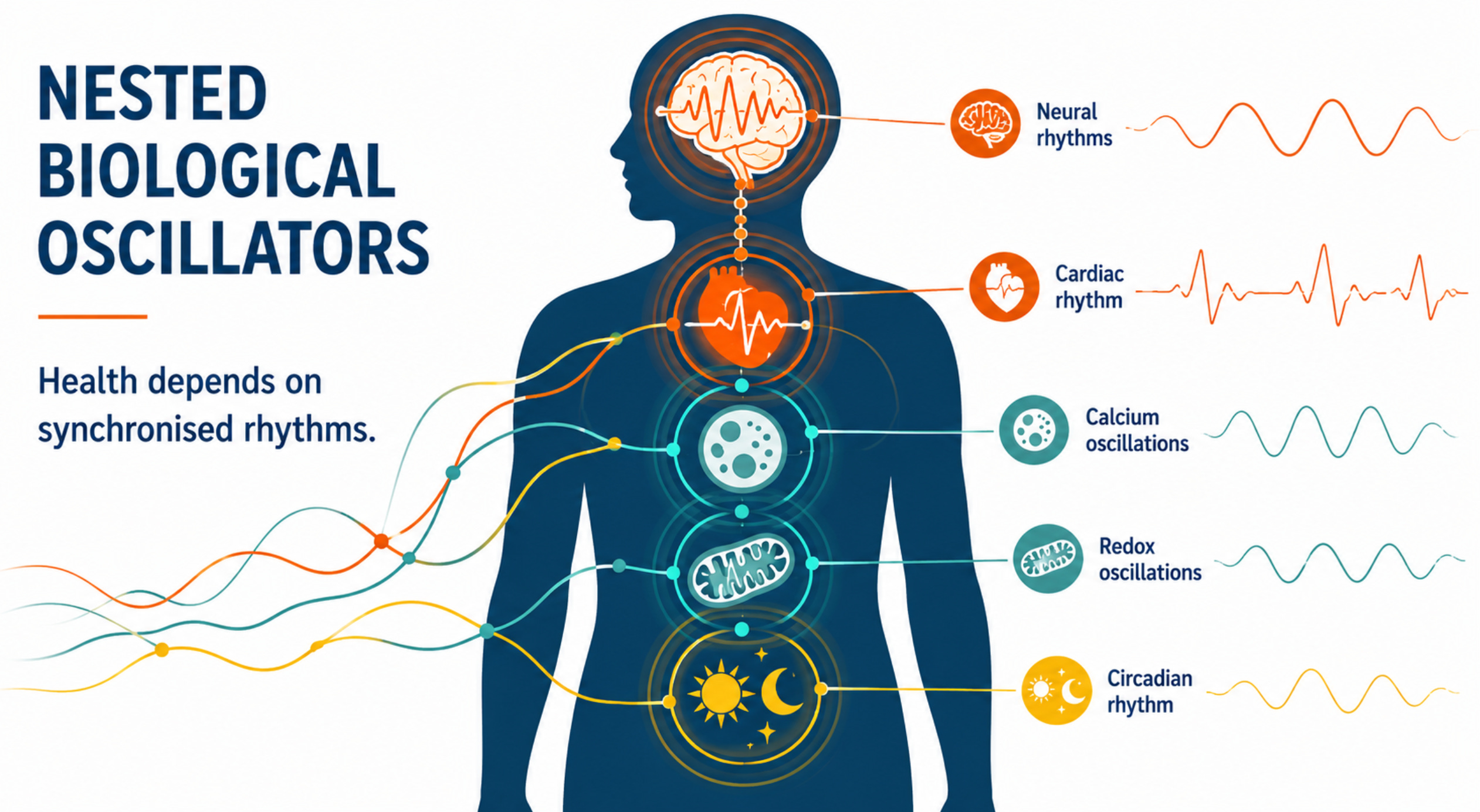
Health depends not simply on the presence of these signals but on the precision of their timing, phase relationships, and synchronisation. Cells with exceptionally high electrical activity and metabolic demand therefore depend most heavily on maintaining this temporal organisation.
The empirical evidence supports this vulnerability. Oxidative stress (excess reactive oxygen species damaging lipids, proteins, and DNA) is among the most consistently replicated findings across in vitro, in vivo, and epidemiological studies of RF-EMF and ELF exposure, observed at field intensities well below thermal thresholds. (Yakymenko et al. 2016; Weller et al. 2025) Notably, the tissues implicated in long-term animal studies are not arbitrary: chronic low-intensity RF exposure has been linked to cardiac Schwannomas and brain gliomas, tumours arising in precisely the energy-intensive, mitochondria-rich neural tissues that this paper identifies as most vulnerable, and which the NTP noted share functionally related cell lineages. (Falcioni et al. 2018; NTP 2018)
Neurons, cardiac muscle, reproductive tissues and other energy-intensive cell types are not merely rich in ion channels and mitochondria; they are among the most highly oscillatory tissues in the body, making them potentially more sensitive to disturbances in biological timing. (Krylov 2023;2026; Damulewicz et al 2025)
Recent experimental evidence supports this directly: frequency-dependent amplification of mitochondrial ROS oscillations by weak ELF magnetic fields has been demonstrated in live cancer cell lines, with the effect dependent on mitochondrial network topology and intrinsic oscillatory state, precisely the properties that distinguish high-demand tissues. (Zandieh et al. 2025)
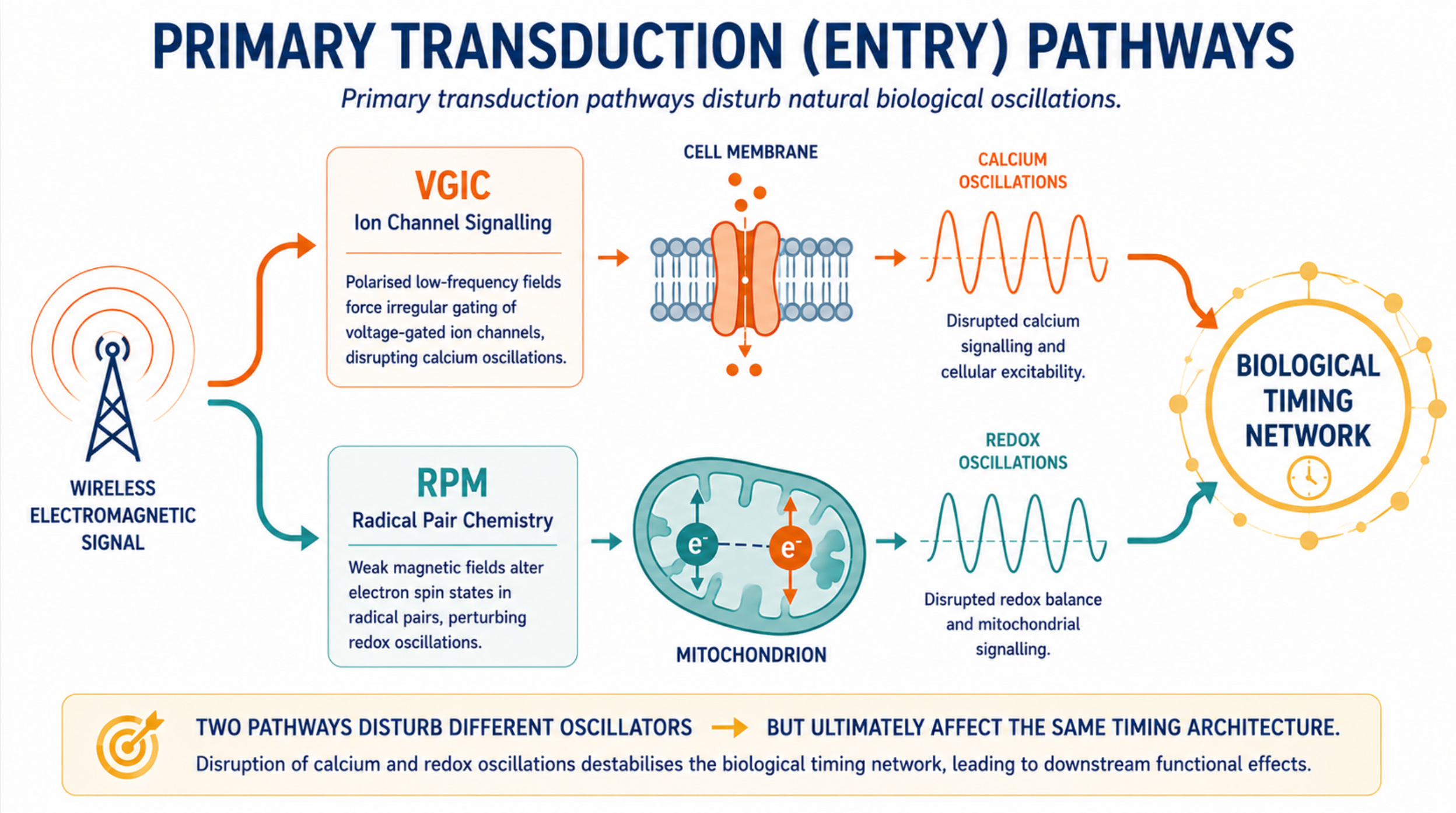
The primary entry point is voltage-gated ion channel dysfunction, described quantitatively by the Ion Forced Oscillation–Voltage-Gated Ion Channel (IFO–VGIC) framework: polarized, coherent, low-frequency field components force irregular gating of voltage sensors, disrupting calcium homeostasis and triggering ROS overproduction (Panagopoulos et al. 2015; 2021; 2025)
However, a second pathway operates in parallel and through different physics entirely, the radical pair mechanism, acting via the magnetic field component of the same signal on flavin-superoxide pairs at the mitochondrial membrane. A third layer, largely invisible until recent quantum biology work, concerns the DNA as a chiral quantum wire whose charge-transport and spin-selective properties are biologically active and structurally coupled to the epigenetic and repair machinery.
This paper examines how all three pathways relate to one another, why they converge on the same tissues, and what the emerging molecular evidence, including the Cyb5b gene-switch finding, the CISS characterisation of DNA, and the Talbi et al (2025) modelling study on GHz carrier frequencies, clarifies - and what it leaves open.
This Substack article is part of a PSGR New Zealand series:
EMF Sensitive? You May Not Be the Outlier You Think You Are.
The EMF Problem: RF Radiation Governance Without Democratic Risk Assessment.
EMF Risk Blindspot: The role of modulation in signal interaction.
The Wired and Unwell Cascade - brain and nervous system harm.
THE IFO–VGIC MECHANISM: CLASSICAL PHYSICS AT THE PLASMA MEMBRANE
‘Significant scientific evidence shows that the bioactivity of WC EMFs is mainly due to their ELF/ULF components and that RF/microwave carrier signals alone, without modulation, pulsing, and variability, do not usually induce biological effects other than heating at adequately high intensities and frequencies (Bawin et al. 1975; 1978; Blackman et al. 1982; Frei et al. 1988; Walleczek 1992; Bolshakov and Alekseev 1992; Goodman et al. 1995; Penafel et al. 1997; Creasey and Goldberg 2001; Huber et al. 2002; Betti et al. 2004; Goldsworthy 2006; Höytö et al. 2008; Franzellitti et al. 2010; Campisi et al. 2010; Mohammed et al. 2013; Panagopoulos 2019). As summarized by Goldsworthy (2006), “it is widely accepted that continuous unmodulated radio waves are of too high a frequency to give biological effects but they do become effective when pulsed or amplitude modulated at a low frequency”.
The Ion Forced Oscillation–Voltage-Gated Ion Channel (IFO–VGIC) mechanism, developed quantitatively by Panagopoulos and colleagues, describes how polarized, coherent, low-frequency fields displace mobile ions within channel pores, forcing the voltage-sensing S4 helices of VGICs to gate irregularly. The result is disrupted calcium homeostasis, which triggers reactive oxygen species (ROS) overproduction through the mitochondrial electron transport chain and NADPH oxidases. Because virtually every cell in the body carries voltage-gated channels, this mechanism operates universally, but its effects concentrate where VGIC density is highest.
The mechanism reflects classical physics principles: Coulomb forces on charged ions, displacement of sensor helices, downstream ionic imbalance. It is also exquisitely frequency- and waveform-dependent, responding specifically to the extremely low frequency (ELF) and ultra-low frequency (ULF) components of modern wireless signals, the pulsing frame rates, modulation envelopes, and chaotic amplitude variability that all digital wireless systems embed in their GHz carrier waves. These ELF and modulation components are the likely mechanism – the biologically relevant component of the signal.
[1] THE RADICAL PAIR MECHANISM: A PARALLEL QUANTUM PATHWAY AT THE MITOCHONDRIAL MEMBRANE
The radical pair mechanism (RPM) is the same quantum process that underlies the avian magnetic compass, demonstrated in cryptochrome proteins in birds’ eyes, where weak magnetic fields alter the singlet-triplet interconversion of electron spin states in radical pairs, changing the ratio of chemical products. In the context of EMF biology, the RPM is relevant not because of its role in navigation but because of where it operates in human cells: within the flavin-containing enzymes of the mitochondrial electron transport chain and NADPH oxidases, where flavin-superoxide radical pairs form continuously during normal oxidative metabolism. (Henshaw and Philips 2024)
What makes this mechanistically significant is that the RPM acts through the magnetic field component of an electromagnetic signal, not the electric field component that drives VGIC dysfunction. The two pathways are therefore mechanistically separate in their physics: they pick up different components of the same wireless signal simultaneously, act on different cellular compartments, and operate through entirely different physical principles, one classical, one quantum. A single exposure event may engage both simultaneously.
Radical-pair reactions may not themselves be the principal biological amplifier. Rather, they may act as triggers within existing biochemical oscillators, particularly mitochondrial redox oscillators, whose intrinsic feedback loops amplify small magnetic perturbations into larger physiological responses. (Krylov 2023;2026) Under this framework, the biological significance of RPM lies not in the magnitude of the initial perturbation but in its ability to alter the timing, synchronisation, or phase relationships of oscillatory redox networks that are already operating within the cell.
This shifts the mechanistic question from energy deposition to timing disruption: the relevant issue is not how much energy is transferred, but whether weak magnetic perturbations can alter the behaviour of biological oscillators that are already close to critical transition points.
Current evidence suggests the RPM does not stand alone as an independent primary mechanism in the context of wireless EMF biology. Talbi et al. (2025) paper examined whether GHz carrier waves can coherently influence radical-pair chemistry before thermal decoherence destroys spin coherence. The answer suggests no, as the carrier oscillates far too rapidly. A 2.4 GHz WiFi carrier completes a full oscillation in 0.4 nanoseconds, whereas spin dynamics in flavin-superoxide radical pairs operate on nanosecond-to-microsecond timescales. The magnetic field component at environmental intensities seems unlikely to accumulate coherent effects across multiple cycles at these frequencies. The GHz carrier itself is unlikely to be the primary driver of RPM-mediated biological responses.
Instead, the mechanistic literature points to modulation structure and low-frequency components as potentially important determinants of biological interaction. At ELF frequencies such as the 217 Hz frame rate of 2G GSM, the 100 Hz pulsing of DECT phones, or the 10 Hz beacon interval of WiFi, the relevant timescales shift from sub-nanosecond carrier oscillations to millisecond-scale modulation patterns. These timescales are substantially closer to those associated with biological spin dynamics and radical-pair processes than the GHz carrier itself. (Henshaw and Philips 2024; Talbi et al 2025) Viewed from this perspective, the RPM is therefore not an alternative to the VGIC framework; but a parallel consequence of the same upstream ELF forcing function, acting through different physics on a different cellular compartment.
Tissue specificity: why VGIC and RPM vulnerability maps coincide
The potential influence of RPM would be expected to increase in tissues with greater densities of flavin-containing redox systems. This creates a tissue-specific sensitivity map that closely mirrors the VGIC vulnerability map.
Neurons carry extraordinary mitochondrial density (Mahadevan et al 2021; Trigo et al 2022) and high NADPH oxidase activity (Belarbi et al 2017; Tu et al 2023). Spermatogenic cells, particularly Leydig and Sertoli cells, have high mitochondrial densities reflecting the energetic demands of testosterone biosynthesis (Andric and Kostic 2019; Zirkin and Papadopoulos 2018). Cardiac muscle is composed of approximately 30-35% mitochondria by cell volume (Brown et al 2017; Piquereau et al 2013). Ovarian granulosa cells sustain high flavin-dependent enzymatic activity during steroidogenesis. These tissues rank among the strongest candidates for VGIC and RPM sensitivity criteria simultaneously (Alberico and Woods 2022; Inaoka et al 2008). The convergence reflects that both mechanisms are driven by the same ELF envelope and amplified by the similar metabolic characteristics.
Molecular convergence: the Cyb5b gene switch and the limits of the GHz carrier
A 2026 paper in Cell by Kim and colleagues (2026) working toward a therapeutic gene-editing tool, identified cytochrome b5 type B (Cyb5b), an outer mitochondrial membrane redox protein, as an essential mediator of an EMF-inducible gene switch discovered through an unbiased CRISPR-Cas9 screen. Critically, the switch was activated by rhythmic oscillatory calcium dynamics rather than by a generic rise in bulk calcium concentration. This provides direct experimental evidence that cells can respond to the pattern of calcium oscillations as information, rather than merely to the overall quantity of calcium present. (Kim et al 2026)
Two important caveats deserve emphasis. First, the Kim experiment used a clean, uniform 60 Hz field, the most favourable possible waveform for biological entrainment. The extrapolation to chaotic, polychromatic wireless signals (complex wireless signals containing multiple frequencies and modulation patterns) disrupting rather than activating calcium oscillation programs is mechanistically plausible. However, this is yet to be directly tested through the Cyb5b pathway. Second, the paper was designed to engineer a therapeutic gene switch, not to study adverse EMF effects, and should not be overstated beyond what it demonstrates.
The Kim et al. paper identified Cyb5b as essential to an EMF-inducible gene switch activated by rhythmic calcium dynamics, a transducer of calcium pattern. Cyb5b sits at a location (the outer mitochondrial membrane) where both VGIC-driven calcium dysrhythmia and RPM-mediated flavin-superoxide perturbation would likely converge. Whether it reads either or both in the context of wireless EMF exposure remains untested. (Kim et al 2026)
The Talbi et al (2025) paper demonstrated that the GHz carrier wave cannot drive RPM chemistry. Talbi et al. proved nothing about whether ELF modulation envelopes degrade calcium timing fidelity (the ability of cells to maintain precise biological timing signals) through VGIC dysfunction, nor anything about Cyb5b-mediated oscillatory disruption. Their model addressed only the magnetic component of GHz carrier waves acting on a simplified radical pair system, a legitimate and important result, but not a complete description of how EMF biology operates in living cells.
[2] DNA AS A CHIRAL QUANTUM WIRE: BEYOND THE FRACTAL ANTENNA HYPOTHESIS
The original Blank-Goodman (2011) hypothesis has, by 2026, been substantially deepened and partially transformed. Their argument identified a plausible mechanism, EMF interaction with delocalised π electrons causing local charging and strand separation, but could not specify how this would account for the observed broad frequency response, nor provide the quantum physical grounding for why this interaction would occur at the extremely low field strengths reported. Since that time the underlying physics has been developed independently. DNA is an electronically active, spin-selective biological conductor embedded within redox, repair and epigenetic networks.
The literature suggests a shift from conceptualising DNA as a literal broadband antenna, to considering that DNA functions as a dynamic charge-transport network whose electronic properties are biologically active and structurally regulated. Whether this arises from environmental electromagnetic fields that amplify or fragment these charges and specifically perturb that system is an important question.
Quantum charge transport in DNA: evidence for biological relevance
The longstanding debate over whether DNA conducts charge, which ranged from reports of metallic-like conduction to semiconductor behavior to insulation, depending on experimental conditions, has narrowed as a consequence of the Ulm group’s theoretical and experimental program from 2023 to 2025. Mirko Rossini and colleagues (2025) directly addressed the question Blank and Goodman left open: does quantum charge diffusion survive in physiological conditions, or does environmental noise destroy coherence?
By modelling quantum diffusion in DNA-inspired two-dimensional tight-binding lattices with both intrinsic and environmental fluctuations, and using atomistic parametrisation, the group found a complex network of charge transport pathways. Long-range quantum phenomena depend on the carrier type (electrons or holes), base sequence, and noise/disorder characteristics. Notably, spatially correlated low-frequency fluctuations can sustain coherent charge transfer across several bases, and moderate vibrational noise can enhance rather than suppress quantum coherence by facilitating tunnelling effects. The authors caution that results apply to short oligomers and that the appropriate coupling scheme for realistic DNA environments remains an open question.
Sequence-dependent charge transport implies that different genomic regions may differ in their susceptibility to charge-transfer perturbations. This could translate into differential sensitivity to externally applied electromagnetic fields, however this is yet to be confirmed.
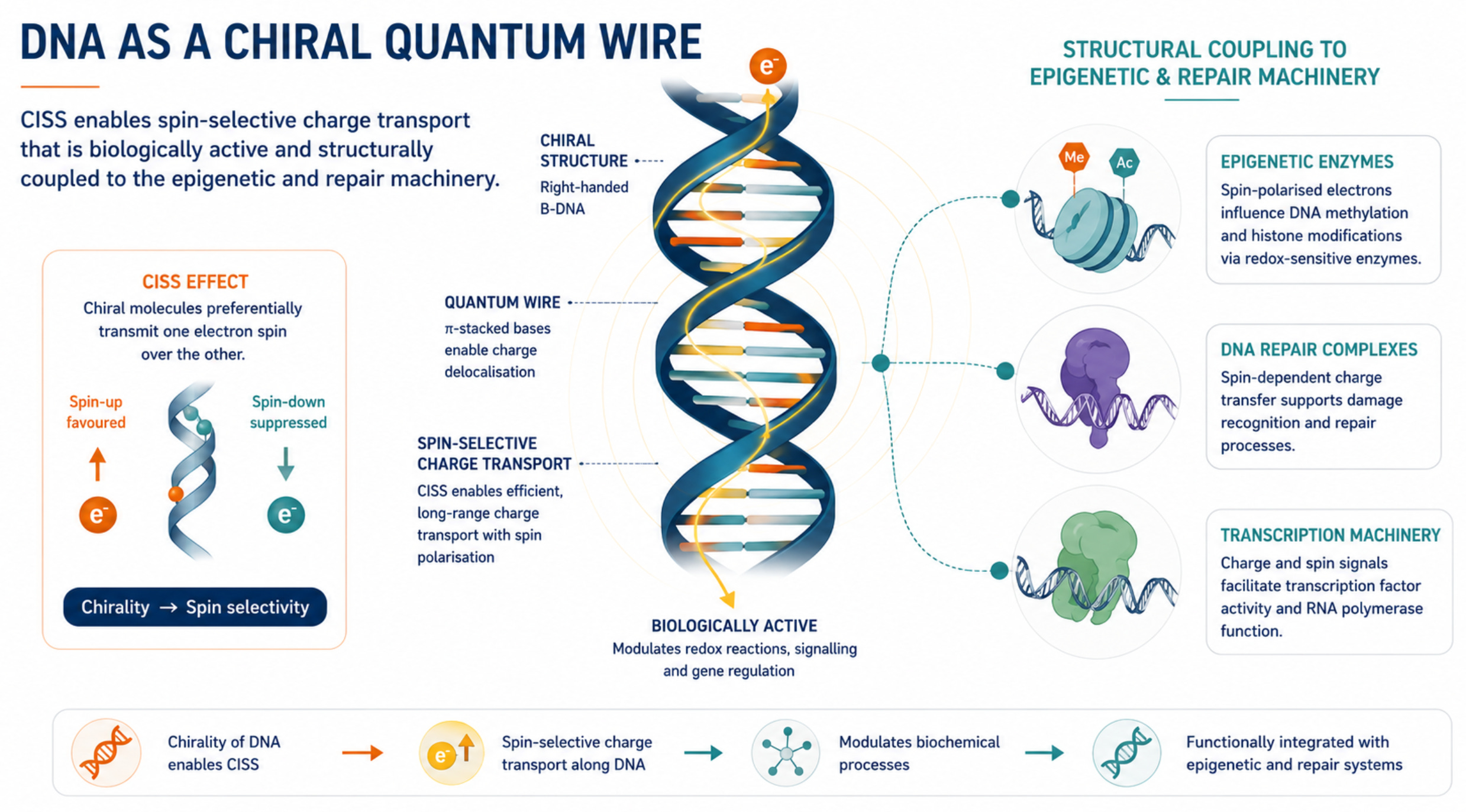
Chirality-induced spin selectivity: DNA as a spin filter
The most significant development since Blank and Goodman is Chirality-Induced Spin Selectivity (CISS), which they did not know about or discuss. CISS was discovered to be active in DNA in 2011, the same year as the fractal antenna paper (Xie et al 2011). The CISS effect refers to ‘the innate ability of chiral materials to act as spin filters for electron transport; however, more recent experiments demonstrate that displacement currents arising from charge polarization of chiral molecules lead to spin polarization without the need for net charge flow.’ (Bloom et al 2024)
The B-form DNA double helix is a right-handed chiral molecule. CISS means that when electrons are transmitted through this chiral structure, they become spin-polarized. i.e., their quantum spin state is filtered by the helical geometry itself, without requiring any magnetic centers or strong spin-orbit coupling. Spin polarization values of approximately 30-60% for DNA hairpins have been measured experimentally using time-resolved EPR under controlled in vitro conditions, with the contribution appearing to increase with applied magnetic field strength. (Siebert et al 2023; Latawiec et al 2025)
The Siebert et al. 2023 Clinical Epigenetics paper from Ulm University (2023), published by a team including quantum physicists Martin Plenio and Fedor Jelezko, proposes a quantum physics layer of epigenetics based on charge transfer and CISS in DNA. Their central argument is that coherent charge transfer along overlapping π-orbitals of DNA bases, combined with CISS-mediated spin selectivity, may act as another regulatory layer, another epigenetic mechanism operating in parallel with classical chemical mechanisms like methylation and histone modification. Critically, they show that charge transfer is not prevented or grossly hindered by DNA methylation or binding to histones, and that if protein binding to DNA does not affect proper base stacking, it preserves coherent charge transfer. This means the quantum layer operates through and alongside the chemical layer.
The guanine finding is especially relevant because guanine acts as a preferred trap for positive charges (‘holes’) moving through DNA. Of the four DNA bases, guanine is the most easily oxidised, meaning that charge tends to accumulate there. This is important because the cytosine paired with guanine is also the primary site of epigenetic methylation at CpG regions, which play a major role in regulating gene activity. As a result, the locations most important for DNA charge transport sit immediately alongside some of the most important sites for epigenetic regulation, creating a plausible interface between quantum charge-transfer processes and the classical machinery that controls gene expression.
In a large review by Weller et al, (2025) two most frequently measured biomarkers were on the same site that quantum biology identifies as the most electronically vulnerable: 8-oxo-7,8-dihydro-2′-deoxyguanosine (8-oxo-dG) and 8-hydroxy-2′-deoxyguanosine (8-OHdG). These are established markers of oxidative DNA damage and risk factors for cancer and neurodegenerative disease (Wang et al 2025).
The CISS/quantum transport framework specifically predicts vulnerability at base-level chemistry, oxidative modification of guanine’s π-electron system, rather than at the sugar-phosphate backbone. DNA base damage emerged as one of the most consistently reported findings in the reviewed literature. Weller et al.’s review found that the highest proportion of statistically significant effects across all study types: 92% of 12 in vitro studies, 80% of 20 in vivo studies, and 100% of 4 epidemiological studies was in the form of DNA base damage. Weller et al, (2025)
Then in August 2025, a PNAS paper from Wasielewski’s group at Northwestern provided direct experimental evidence of CISS in isolated DNA hairpins using time-resolved electron paramagnetic resonance, showing that CISS significantly influences the spin dynamics of radical pairs produced by photoexcitation, imparting partial triplet character to the spin-correlated radical pair. The CISS contribution was found to increase with applied magnetic field strength rather than with DNA length, consistent with hole delocalization over guanine tracts. (Latawiec et al 2025)
What remains of the fractal antenna: CISS, oxidative feedback, and open questions
The original Blank-Goodman argument proposed that EMF interacts with delocalised π electrons in DNA bases, causing local charging and strand separation. The paper did not explain why this interaction would occur across such a broad and featureless frequency range without thermal energy, or to ground it in the quantum physics of the molecule.
Unlike a radio antenna absorbing energy from an external field and re-emitting, the DNA helix mechanism is subtler and more biologically integrated. What the CISS and quantum charge transport work reveals is that DNA is a spin-selective quantum wire embedded in a biological system where charge transport is functionally coupled to repair, replication, and epigenetic marking. In principle, spin-selective transport systems can exhibit magnetic-field sensitivity. Whether DNA-associated CISS processes remain sufficiently coherent and magnetically responsive in intact chromatin to be perturbed by weak environmental fields remains an open question.
The oxidatively damaged DNA work is critical here: a 2019 study in JACS found enhancement in spin-dependent transport through oxidatively damaged DNA.(Mishra et al 2019) This means the CISS effect is not just a property of intact DNA, it is altered by oxidative damage in a way that changes the charge transport channel from the base stack to the sugar-phosphate backbone, which is less conductive but more spin-selective. If EMF-induced ROS (through the VGIC and mitochondrial pathways) causes oxidative damage, that damage changes the charge transport properties of DNA, which then alters the CISS-dependent spin chemistry, which feeds back into further damage at guanine sites. This suggests a positive feedback loop between oxidative damage and quantum transport perturbation that Blank and Goodman could not have articulated in 2011.
Environmental noise as a feature, not a bug
In physics, ‘noise’ refers to the random jostling and fluctuations that surround any molecule in a living cell, the thermal vibrations of water, the movement of ions, the constant molecular bustle of the biological environment, and the conventional assumption has been that this noise would simply scramble any delicate quantum effects in DNA before they could do anything useful.
The Rossini et al. finding that a certain degree of environmental noise supports rather than simply destroys coherent charge propagation in short DNA sequences addresses a central physics objection to the fractal antenna idea, that thermal decoherence at physiological temperature would collapse any quantum effects.
Their model shows that proximity-correlated low-frequency fluctuations can sustain and even enhance quantum coherence along the π-stack, a result analogous to noise-assisted transport in photosynthesis. It remains an open question whether external electromagnetic fields, the ELF components of wireless signals, would interact with this existing noise landscape in biologically significant ways. The biological noise modelled by Rossini et al. is intrinsic (thermal, backbone vibrations, solvent), and no study has yet examined whether extrinsic RF or ELF fields would similarly assist or perturb quantum charge diffusion along the π-stack in living cells. (Rossini et al. 2025) The amplitude-window (a non-linear dose-response with a peak effect at intermediate field strengths) and the IFO-VGIC framework both suggest that biological sensitivity to field strength is non-monotonic, which would be consistent with a noise-assisted quantum transport system. (Goldsworthy 2012; Panagopoulos 2010)
This work provides a plausible mechanism whereby wireless EMF exposure at environmental intensities perturbs CISS or quantum charge transport in living cells. The CISS research has been done with purified DNA on surfaces or in solution, not in intact chromatin in nuclei. The noise-assisted transport models are theoretical. The epigenetic quantum layer is a hypothesis in Clinical Epigenetics, not a demonstrated mechanism with causal chains mapped in vivo. And the Blank-Goodman proposal itself, that DNA’s broadband frequency response explains the wide range of EMF biological effects, remains descriptively plausible but not quantitatively predictive in the way Panagopoulos’ equations are.
Synthesis: one upstream driver, three transduction pathways
Blank and Goodman identified the right molecule but lacked the physics to say why. Fifteen years of quantum biology has now supplied it: DNA as a chiral quantum wire in which coherent charge transport along the π-stack is spin-selective due to CISS, sequence-dependent in its energy landscape with guanine as the preferred charge trap, coupled to the epigenetic machinery at CpG sites, noise-assisted in its transport dynamics, and altered in both conductance and spin selectivity by oxidative damage. Although no experimental study has yet shown that environmental ELF fields or wireless modulation patterns directly perturb DNA CISS-mediated charge transport in living cells, the mechanistic literature provides a plausible framework for asking that question.
[3] SECONDARY, BRIDGING MECHANISMS
Other mechanisms frequently discussed in the non-thermal EMF literature include membrane lipid organisation, cytoskeletal signalling, blood-brain barrier permeability, immune and inflammatory activation, heat-shock responses, and disruption of circadian and melatonin regulation through cryptochrome-dependent pathways. These may all contribute to the observed biological picture, but they are generally better understood as downstream, tissue-specific, or bridging mechanisms rather than primary transduction pathways. In most proposed models, the initial interaction occurs either through altered ion-channel activity and calcium signalling, or through magnetic-field effects on radical-pair chemistry. (Belpomme and Irigaray 2021; Jangid et al 2025; Leszczynski 2022; 2025; Panagopoulos et al. 2025)
The resulting disturbances then propagate through established biological networks involving oxidative stress, mitochondrial dysfunction, inflammatory signalling, endocrine regulation, DNA repair, and circadian control. In this sense, many apparently distinct observations in the literature may represent different manifestations of a common upstream disturbance rather than entirely separate mechanisms. The growing literature on cryptochrome, magnetoreception, circadian biology, and melatonin regulation is particularly relevant because it links radical-pair chemistry to whole-organism timing systems, providing a plausible route by which small molecular perturbations could influence sleep, hormonal rhythms, immune function, and cellular repair processes. (Damulewicz et al 2025; Eskandani and Zibaii 2024; Martel et al. 2023; Pall 2016; Panagopoulos et al. 2025)
A particularly important recent development is the proposal that mitochondrial reactive oxygen species (ROS) oscillations may act as a biological amplification system. One of the major objections to weak-field effects has always been the so-called ‘kT problem’: the magnetic energies involved are many orders of magnitude smaller than thermal energy and therefore appear incapable of producing biologically meaningful change. Zandieh and colleagues (2025) addressed this problem directly by proposing that weak magnetic perturbations arising from radical-pair chemistry need not produce large direct effects. Instead, they may act on pre-existing mitochondrial redox oscillators that are already capable of generating large-scale cellular dynamics.
Rather than behaving as static generators of ROS, mitochondria can function as coupled oscillatory networks exhibiting intrinsic redox rhythms, ROS-induced ROS-release (RIRR), and feedback-driven synchronisation. This concept builds on earlier work by Aon, Cortassa and colleagues demonstrating that mitochondrial populations can undergo coordinated oscillatory behaviour and phase transitions under conditions of oxidative stress. This work has since developed into a broader systems-biology framework in which mitochondria are understood as dynamic components of nested biological networks spanning scales from molecular reactions to whole-organism physiology. (Aon and Cortassa 2026)
In such systems, a very small perturbation introduced at the radical-pair level may be amplified through resonance, phase-shifting, the triggering of oscillatory behaviour in systems poised near a threshold, or the synchronisation of pre-existing oscillatory patches into a larger coherent network.
The significance of the Zandieh model is that it reframes weak-field interactions as a problem of timing rather than energy transfer. In their formulation, magnetic-field-induced changes in radical-pair chemistry within mitochondrial Complex III slightly alter superoxide production. Under appropriate physiological conditions, these small changes can interact with intrinsic mitochondrial oscillators and produce disproportionately large downstream effects on membrane potential, calcium signalling, gene expression, and cellular stress responses. The model predicts frequency-dependent biological windows in the hertz and sub-hertz range and shifts attention away from total energy deposition toward the interaction of weak fields with pre-existing biological oscillators (Zandieh et al., 2025).
Weak-field effects can therefore be understood within a broader systems-biology framework in which living organisms are organised as hierarchies of nested oscillators, spanning scales from molecular reactions and mitochondrial redox cycles to neuronal networks, endocrine rhythms, and circadian clocks.
In this interpretation, amplification arises not because the field supplies biologically significant energy, but because it perturbs the timing and synchronisation of endogenous oscillatory processes that are already present within the cell. While this remains an emerging area of research and requires substantial replication, it provides one of the most coherent theoretical explanations yet proposed for how extremely weak ELF magnetic fields could influence cellular function without invoking thermal mechanisms. (Krylov 2023;2026; Zandieh et al. 2025)
Importantly, this framework predicts frequency-dependent biological windows in the hertz and sub-hertz range and shifts attention away from total energy deposition toward the interaction of weak fields with pre-existing biological oscillators. (Zandieh et al. 2025) While this remains an emerging area of research and requires substantial replication, it provides one of the most plausible theoretical explanations yet proposed for how extremely weak ELF magnetic fields could influence cellular function without invoking thermal mechanisms.
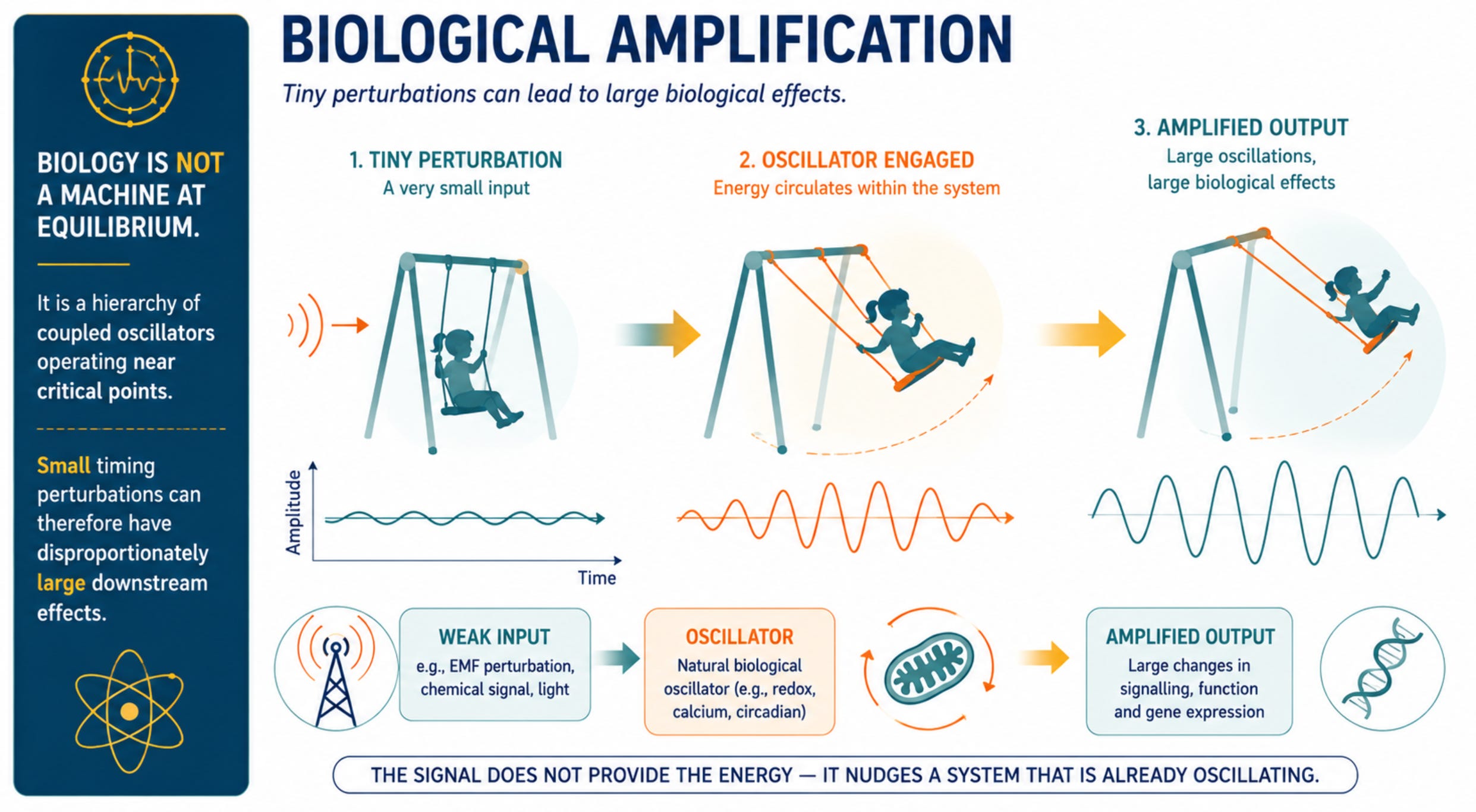
CONCLUSION
A single upstream physical feature, ELF/ULF variability embedded within modern wireless signals, emerges as a credible common driver of biological effects. Living systems are organised around nested oscillators operating across many timescales, including membrane potentials, calcium signals, mitochondrial redox rhythms, neuronal synchronisation, hormone signalling rhythms, and circadian clocks. Health depends not simply on the presence of these signals, but on the precision of their timing, phase relationships, and synchronisation.
ELF/ULF variability couples into living biology through at least two independent transduction pathways simultaneously. The VGIC pathway acts at the plasma membrane via classical physics, disrupting ionic balance and calcium homeostasis. The RPM pathway acts at the mitochondrial inner membrane and NADPH oxidases via quantum spin chemistry, perturbing the superoxide/hydrogen peroxide ratio. Both converge on ROS overproduction, oxidative stress, and, in chronically exposed tissues, DNA damage and cellular dysfunction.
Although the VGIC and RPM pathways operate through different physics, they converge on the same biological property: the regulation of oscillatory processes. The VGIC pathway primarily perturbs calcium oscillations and membrane excitability, whereas the RPM pathway perturbs mitochondrial redox oscillations and reactive oxygen species dynamics. Both therefore act not by supplying large amounts of energy, but by altering the timing and synchronisation of biological systems that are already oscillating.
The reason ELF modulation components rather than the GHz carrier are the more plausible bioactive signal is mechanistically grounded: biological charge-transfer and radical-pair processes operate over microsecond-to-second timescales, placing them in register with pulsed structure, intermittency, and beat-frequency phenomena, rather than with the carrier itself.
Cyb5b sits at the outer mitochondrial membrane, a location where VGIC-driven calcium dysrhythmia and RPM-mediated flavin-superoxide perturbation would converge. Whether it transduces either or both signals under wireless EMF exposure is not yet known; but if it does, it would represent a single molecular node through which classical and quantum pathways converge to influence gene regulation.
DNA is not a passive downstream target of this oxidative cascade. The ROS generated through the VGIC and RPM pathways causes oxidative damage that alters DNA charge transport and CISS-dependent behaviour, raising the possibility that DNA participates actively in the propagation and amplification of oxidative injury, in a manner that Blank and Goodman anticipated but could not mechanistically specify.
Small perturbations introduced into these systems may then be amplified through the feedback architecture of living cells, particularly within mitochondrial redox networks and calcium signalling pathways.
Together, these observations support a credible and increasingly well-developed hypothesis: that chronic exposure to complex, non-native ELF envelopes may degrade biological timing fidelity in tissues that depend most heavily on precise electrical and metabolic coordination. Neurons, cardiac muscle, reproductive tissues, and other highly energy-dependent cell types emerge repeatedly as vulnerable not simply because they contain large numbers of ion channels and mitochondria, but because they are among the most highly oscillatory tissues in the body.
REFERENCES
Alberico HC and Woods DC (2022) Role of Granulosa Cells in the Aging Ovarian Landscape: A Focus on Mitochondrial and Metabolic Function. Front. Physiol. 12:800739. doi: 10.3389/fphys.2021.800739
Andric SA and Kostic TS. Regulation of Leydig cell steroidogenesis: intriguing network of signaling pathways and mitochondrial signalosome. Current Opinion in Endocrine and Metabolic Research, 6:7-20, https://doi.org/10.1016/j.coemr.2019.03.001
Aon MA, Cortassa S. 2026. Systems Biology of Aging, Metabolism, and Mitochondria. Annual Review Biophysics. 55:443-467. https://doi.org/10.1146/annurev-biophys-021424-011852
Belarbi K, Cuvelier E, Destée A, Gressier B, Chartier-Harlin MC. NADPH oxidases in Parkinson’s disease: a systematic review. Mol Neurodegener. 2017 Nov 13;12(1):84. doi: 10.1186/s13024-017-0225-5.
Belpomme D, Irigaray P (2022). Why Electrohypersensitivity and Related Symptoms Are Caused by Non-Ionizing Man-Made Electromagnetic Fields: An Overview and Medical Assessment. Environmental Research. Volume 212, Part A, September 2022, 113374. https://doi.org/10.1016/j.envres.2022.113374
Blank, M., & Goodman, R. (2011). DNA is a fractal antenna in electromagnetic fields. International Journal of Radiation Biology, 87(4), 409–415. https://doi.org/10.3109/09553002.2011.538130
Bloom BP, Paltiel Y, Naaman R, and Waldeck DH. Chemical Reviews 2024 124 (4), 1950-1991, DOI: 10.1021/acs.chemrev.3c00661
Brown, D., Perry, J., Allen, M. et al. Mitochondrial function as a therapeutic target in heart failure. Nat Rev Cardiol 14, 238–250 (2017). https://doi.org/10.1038/nrcardio.2016.203
Damulewicz, M., Gregoris, F., Colaianni, D. et al. Cryptochrome interaction networks across different tissues in Drosophila melanogaster. Biol Direct 20, 114 (2025). https://doi.org/10.1186/s13062-025-00696-x
Eskandani R and Zibaii MI, Unveiling the biological effects of radio-frequency and extremely low frequency electromagnetic fields on the central nervous system performance. BioImpacts. 2024;14(4):30064 doi: 10.34172/bi.2023.30064
Falcioni L, Bua L, Tibaldi E, et al. 2018. Report of final results regarding brain and heart tumors in Sprague-Dawley rats exposed from prenatal life until natural death to mobile phone radiofrequency field representative of a 1.8 GHz GSM base station environmental emission. Environmental Research 165: 496–503. https://doi.org/10.1016/j.envres.2018.01.037
Henshaw, D. L., & Philips, A. (2024). A mechanistic understanding of human magnetoreception validates the phenomenon of electromagnetic hypersensitivity (EHS). International Journal of Radiation Biology, 101(2), 186–204. https://doi.org/10.1080/09553002.2024.2435329
Inaoka, Y., Yazawa, T., Mizutani, T. et al. Regulation of P450 oxidoreductase by gonadotropins in rat ovary and its effect on estrogen production. Reprod Biol Endocrinol 6, 62 (2008). https://doi.org/10.1186/1477-7827-6-62
Jangid P, Rai U, Ahmed S et al. 2025. Non-thermal biological effects of radiofrequency electromagnetic radiation: Mechanistic insights into male reproductive vulnerability in the era of ubiquitous exposure , Reproductive Toxicology, 138:109087, https://doi.org/10.1016/j.reprotox.2025.109087
Kim J, Hwang Y, Kim S et al. Electromagnetic field-inducible in vivo gene switch for remote spatiotemporal control of gene expression, Cell, 2026; 189, 3465-3480.e23 https://doi.org/10.1016/j.cell.2026.03.029
Krylov VV. Molecular Biological Effects of Weak Low-Frequency Magnetic Fields: Current State and Future Prospects. Preprints.org. Posted 26 May 2023, DOI: 10.20944/preprints202305.1930.v1
Krylov VV. Biological effects of weak magnetic fields: can the radical-pair mechanism provide a universal explanation? Biol Rev Camb Philos Soc. 2026 Apr;101(2):893-910. doi: 10.1111/brv.70108.
Latawiec EI, ChiesaA, Qiu Y, Tcyrulnikov NA,et al. (2025) Detecting chirality-induced spin selectivity in chromophore-linked DNA hairpins using photogenerated radical pairs, Proc. Natl. Acad. Sci. U.S.A. 122 (32) e2515120122, https://doi.org/10.1073/pnas.2515120122 (2025).
Leszczynski D. Review of the scientific evidence on the individual sensitivity to electromagnetic fields (EHS). Reviews on Environmental Health. 2022 Sep;37(3):423-450. DOI: 10.1515/reveh-2021-0038. PMID: 34229366.
Leszczynski D, de Vocht F. Editorial: Individual sensitivity to wireless radiation. Front Public Health. 2025 Dec 5;13:1744897. doi: 10.3389/fpubh.2025.1744897.
Mishra S, Poonia VS, Fontanesi C, Naaman R, Fleming AM, Burrows CJ. Effect of Oxidative Damage on Charge and Spin Transport in DNA. J Am Chem Soc. 2019;141(1):123–126. doi:10.1021/jacs.8b12014
Mahadevan HM, Hashemiaghdam A, Ashrafi G, Harbauer A, Mitochondria in Neuronal Health: From Energy Metabolism to Parkinson’s Disease. Adv. Biology2021, 5, 2100663. https://doi.org/10.1002/adbi.202100663
Martel J, Chang SH, Chevalier G et al. Influence of electromagnetic fields on the circadian rhythm: Implications for human health and disease. Biomedical Journal. 46(1):48-59, https://doi.org/10.1016/j.bj.2023.01.003
Melnick, R.L., Moskowitz, J.M. & The International Commission on the Biological Effects of Electromagnetic Fields (ICBE-EMF). Exposure limits to radiofrequency EMF do not account for cancer risk or reproductive toxicity assessed from data in experimental animals. Environ Health 25, 42 (2026). https://doi.org/10.1186/s12940-026-01288-6
Mevissen M, Ducray A, Ward, JM, Kopp-Schneider, McNamee JP, Wood WW, Rivero TM, Straif K. Effects of radiofrequency electromagnetic field exposure on cancer in laboratory animal studies, a systematic review. Environ Int. 2025;199:109482. doi:10.1016/j.envint.2025.109482
National Toxicology Program. 595: NTP Technical Report on the Toxicology and Carcinogenesis Studies in Hsd: Sprague Dawley SD Rats Exposed to Whole-Body Radio Frequency Radiation at a Frequency (900 MHz) and Modulations (GSM and CDMA) Used by Cell Phones. National Toxicology Program, US Department of Health and Human Services. 2018. https://ntp.niehs.nih.gov/sites/default/files/ntp/htdocs/lt_rpts/tr595_508.pdf
Pall, M.L., 2016. Microwave frequency electromagnetic fields (EMFs) produce wide- spread neuropsychiatric effects including depression. J. Chem. Neuroanat. 75 (Pt B), 43–51. http://dx.doi.org/10.1016/j.jchemneu.2015.08.001.
Panagopoulos D, Margaritis L. (2010a) The identification of an intensity “window” on the bioeffects of mobile telephony radiation. Int J Radiat Biol 86(5):358–366. DOI: 10.3109/09553000903567979.
Panagopoulos DJ, Johansson O, Carlo GL. Polarization: a key difference between man-made and natural electromagnetic fields, in regard to biological activity. Sci Rep. (2015) 5:14914. doi: 10.1038/srep14914
Panagopoulos DJ, Karabarbounis A, Yakymenko I, Chrousos GP. Human-made electromagnetic fields: ion forced-oscillation and voltage-gated ion channel dysfunction, oxidative stress and DNA damage. Int J Oncol. (2021) 59:92. doi:10.3892/ijo.2021.5272
Panagopoulos DJ. Electromagnetic fields of wireless communications: biological and health effects. 1st ed. Boca Raton: CRC Press (2023).
Panagopoulos DJ, Yakymenko I, De Iuliis GN, Chrousos GP. A comprehensive mechanism of biological and health effects of anthropogenic extremely low frequency and wireless communication electromagnetic fields. Front Public Health. 2025 Jun 4;13:1585441. doi: 10.3389/fpubh.2025.1585441.
Piquereau J, Caffin F, Novotova M, Lemaire C, Veksler V, Garnier A, Ventura-Clapier R, Joubert F. Mitochondrial dynamics in the adult cardiomyocytes: which roles for a highly specialized cell? Front Physiol. 2013 May 10;4:102. doi: 10.3389/fphys.2013.00102
Rossini M, Ammerpohl O, Siebert R, Ankerhold J. Effects of environmental noise on quantum charge diffusion in DNA sequences. Sci Rep. 2025 May 20;15(1):17543. doi: 10.1038/s41598-025-02819-w.
Siebert, R., Ammerpohl, O., Rossini, M. et al. A quantum physics layer of epigenetics: a hypothesis deduced from charge transfer and chirality-induced spin selectivity of DNA. Clin Epigenet 15, 145 (2023). https://doi.org/10.1186/s13148-023-01560-3
Talbi O, Zadeh-Haghighi H and Simon C (2025) The radical pair mechanism cannot explain telecommunication frequency effects on reactive oxygen species. Front. Quantum Sci. Technol. 4:1544473. doi: 10.3389/frqst.2025.1544473
Trigo D, Avelar C, Fernandes M, Sá J, da Cruz E Silva O. Mitochondria, energy, and metabolism in neuronal health and disease. FEBS Lett. 2022 May;596(9):1095-1110. doi: 10.1002/1873-3468.14298.
Tu D, Velagapudi R, Gao Y, Hong JS, Zhou H, Gao HM. Activation of neuronal NADPH oxidase NOX2 promotes inflammatory neurodegeneration. Free Radic Biol Med. 2023 May 1;200:47-58. doi: 10.1016/j.freeradbiomed.2023.03.001.
Wang, J., Li, C., Han, J. et al. Reassessing the roles of oxidative DNA base lesion 8-oxoGua and repair enzyme OGG1 in tumorigenesis. J Biomed Sci 32, 1 (2025). https://doi.org/10.1186/s12929-024-01093-8
Weller SG, McCredden JE, Leach V, Chu C and Lam AK (2025) A scoping review and evidence map of radiofrequency field exposure and genotoxicity: assessing in vivo, in vitro, and epidemiological data. Front. Public Health 13:1613353. doi: 10.3389/fpubh.2025.1613353
Xie Z, Markus TZ, Cohen SR et al. Spin Specific Electron Conduction through DNA Oligomers Nano Lett. 2011, 11, 11, 4652–4655. https://doi.org/10.1021/nl2021637
Yakymenko I, Tsybulin O, Sidorik E, Henshel D, Kyrylenko O, Kyrylenko S. Oxidative mechanisms of biological activity of low-intensity radiofrequency radiation. Electromagn Biol Med. (2016) 35:186–202. doi: 10.3109/15368378.2015.1043557
Zandieh, A., Shariatpanahi, S.P., Ravassipour, A.A. et al. An amplification mechanism for weak ELF magnetic fields quantum-bio effects in cancer cells. Sci Rep 15, 2964 (2025). https://doi.org/10.1038/s41598-025-87235-w
Zirkin BR, Papadopoulos V. Leydig cells: formation, function, and regulation. Biol Reprod. 2018 Jul 1;99(1):101-111. doi: 10.1093/biolre/ioy059. PMID: 29566165